检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
1月13日,我院李光副教授和王义权教授团队在Development杂志上在线发表题为“Cilia-driven asymmetric Hedgehog signalling determines the amphioxus left-right axis by controlling Dand5 expression”的研究论文。该研究深入阐述了纤毛运动和Hh信号在文昌鱼左右不对称建立中的功能及调控机制,为理解胚胎的左右对称如何打破提供了新的见解。这是该团队在《美国科学院院刊》(PNAS,2017)和《发育》(Development,2017)上发表有关文昌鱼左右不对称建立机制研究之后的又一篇相关研究。

我们生活中常见的动物(包括人)大都属于两侧对称动物,因为从外部形态看它们都是左右对称的。然而如果从内部结构看,这些生物却是左右不对称的。比如人(或其他脊椎动物)的心脏总位于身体左侧,而肝脏则总位于身体右侧。这种不对称性在不同个体间十分稳定,说明其建立过程受到严格调控。有趣的是这种不对称性建立于个体发育早期,由左右完全对称的早期胚胎发育而来。那么是什么样的机制将早期胚胎的对称性打破,并驱使胚胎向着左右不对称方向发育的呢?基于脊椎动物模型(如小鼠、斑马鱼等)的研究发现,存在于早期神经胚后端的节点流(nodal flow,由节点处运动纤毛定向转动引起的液流)是打破左右对称的直接驱动力。需要说明的是节点流属于物理性信号。那么这一物理信号又是如何被转化成生物信号,并进而诱导基因(如Cer、Nodal等)不对称表达的呢?过去的研究并未给出明确解释,而只给出了几种可能模型。在这些模型中,two-cilia(双纤毛)模型和morphogen(形态发生素)模型最为流行。
Cer是目前已知的受节点流调控的最早开始左右不对称表达的基因。该团队在前期研究中发现,Hh基因(一种形态发生素)为Cer基因表达所必须,敲除该基因可导致Cer基因无法转录。因而暗示Hh基因位于Cer的上游(Development,2017)。在该研究中他们进一步发现,Hh蛋白及Hh信号靶基因Ptch是左右不对称分布的,且其出现时间和位置与Cer不对称表达时间和位置重叠,而当过激活Hh信号时可引起Cer在左侧异位表达。从而证明了Hh信号的不对称性(右边>左边)可能是导致Cer基因不对称表达的(右边>左边)重要诱因。该研究还显示,抑制胚胎纤毛运动可阻断Hh蛋白的不对称分布,并进而引起胚胎左右不对称发生异常;而去除胚胎纤毛可阻断Hh信号传导,同样导致胚胎左右不对称发生异常。这些结果说明,纤毛运动是导致Hh蛋白不对称分布的驱动力,而纤毛本身又为Hh信号传导所必须。因而该研究不仅论证了纤毛在文昌鱼左右不对称建立中的功能,还揭示了纤毛运动这一物理信号是如何通过影响形态发生素Hh蛋白的分布(生物信号)进而导致基因不对称表达的。该发现统一了双纤毛和形态发生素模型,为理解胚胎的左右对称如何打破提供了新的见解。在该论文发表时,Development杂志还在其The people behind the papers专栏对论文的主要参与者进行了专门采访报道,并通过其官方微信号对该论文进行了宣传。
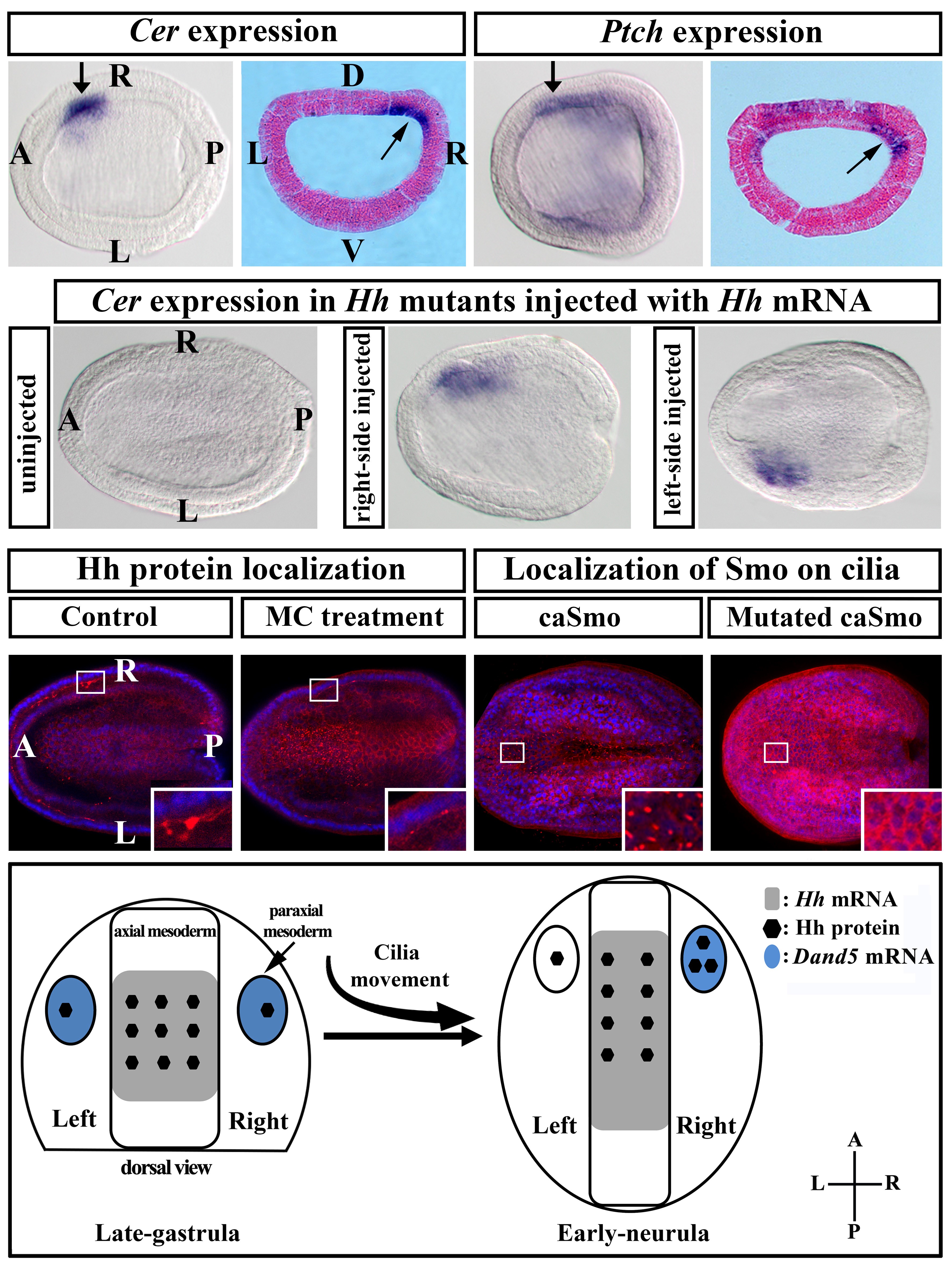
纤毛及Hh信号参与文昌鱼左右不对称建立的机制
伟德BETVlCTOR1946博士生朱鑫为该文第一作者,李光副教授和王义权教授为该文的共同通讯作者。该研究得到了国家自然科学基金委和中央高校基本科研基金的资助。
原文链接:https://dev.biologists.org/content/147/1/dev182469.long
采访链接:https://dev.biologists.org/content/147/1/dev187062.long
微信宣传链接:https://mp.weixin.qq.com/s/cc4dJBCGl0H773UMBfstaA
(伟德BETVlCTOR1946)
